Модель «хищник - жертва» и макроэкономическая модель Гудвина
Рассмотрим биологическую модель «хищник - жертва», в которой один вид является кормом для другого. Эта модель, давно ставшая классической, была построена в первой половине XX в. итальянским математиком В. Вольтерра для объяснения колебаний уловов рыбы в Адриатическом море . В модели предполагается, что число хищников растет до тех пор, пока у них будет достаточно пищи, а увеличение поголовья хищников приводит к уменьшению популяции рыб-жертв. Когда последних становится мало, численность хищников уменьшается. Вследствие этого с некоторого момента начинается рост численности рыб-жертв, который через некоторое время вызывает рост популяции хищников. Цикл замыкается.
Пусть N x (t) и N 2 (t) - численности рыб-жертв и рыб-хищников в момент времени t соответственно. Предположим, что темп прироста численности жертв в условиях отсутствия хищников постоянен, т. е.
где а - положительная постоянная.
Появление хищника должно снизить темп прироста жертв. Будем считать, что это снижение линейно зависит от численности хищника: чем больше хищников, тем меньше темп прироста жертв. Тогда

где т > 0.
Поэтому для динамики численности рыб-жертв получаем:
Составим теперь уравнение, определяющее динамику популяции хищников. Предположим, что их численность в случае отсутствия жертв сокращается (из-за отсутствия корма) с постоянным темпом Ь, т. е.
![]()
Наличие жертв вызывает увеличение темпа прироста хищников. Предположим, что этот прирост линеен, т. е.
![]()
где п> 0.
Тогда для скорости роста рыб-хищников получаем уравнение:
В системе «хищник - жертва» (6.17)-(6.18) снижение скорости роста численности рыб-жертв, вызванное поеданием их хищниками, равно mN x N 2 , т. е. пропорционально числу их встреч с хищником. Увеличение скорости роста численности рыб-хищников, вызванное наличием жертв, равно nN x N 2 , т. е. тоже пропорционально числу встреч жертв и хищников.
Введем безразмерные переменные U = mN 2 /a и V = nN x /b. Динамика переменной U соответствует динамике хищников, а динамика переменной V - динамике жертв. В силу уравнений (6.17) и (6.18) изменение новых переменных определяется системой уравнений:
Предположим, что при t = 0 число особей обоих видов известно, следовательно, известны начальные значения новых переменных?/(0) = U 0 , К(0) = К 0 . Из системы уравнений (6.19) можно найти дифференциальное уравнение для ее фазовых траекторий:

Разделяя переменные этого уравнения, получим:


Рис. 6.10. Построение фазовой траектории ADCBA системы дифференциальных уравнений (6.19)
Отсюда с учетом начальных данных следует:
где константа интегрирования С = b(V Q - InV 0)/a - lnU 0 + U 0 .
На рис. 6.10 показано, как строится линия (6.20) при заданном значении С. Для этого в первой, во второй и в третьей четвертях соответственно строим графики функций х = V - InV, у = (Ь/а)х , у = = InU- U+C.
В силу равенства dx/dV = (V- 1)/У функция х = V- In К, определенная при V > 0, возрастает, если V> 1, и убывает, если V 1. В силу того, что cPx/dV 1 = 1/F 2 > 0, график функции л: = x(V) направлен выпуклостью вниз. Уравнение V= 0 задает вертикальную асимптоту. Наклонных асимптот эта функция не имеет. Поэтому график функции х = х(У) имеет вид кривой, изображенной в первой четверти рис. 6.10.
Аналогичным образом исследуется функция у = InU - U+ С, график которой на рис. 6.10 изображен в третьей четверти.
Если теперь мы разместим на рис. 6.10 во второй четверти график функции у = (Ь/а)х , то в четвертой четверти получим линию, которая связывает переменные U и V. Действительно, взяв точку V t на оси OV, вычисляем с помощью функции х = V - V соответствующее знание х х. После этого, используя функцию у = (Ь/а)х , по полученному значению х { находим у х (вторая четверть на рис. 6.10). Далее с помощью графика функции у = InU - U + С определяем соответствующие значения переменной U (на рис. 6.10 таких значений два - координаты точек М и N). Совокупность всех таких точек (V; U) образует искомую кривую. Из построения следует, что графиком зависимости (6.19) является замкнутая линия, содержащая внутри себя точку Е( 1, 1).
Напомним, что эту кривую мы получили, задав некоторые начальные значения U 0 и V 0 и вычислив по ним константу С. Взяв другие начальные значения, мы получим другую замкнутую линию, не пересекающую первую и также содержащую внутри себя точку Е( 1, 1). Это означает, что семейством траекторий системы (6.19) на фазовой плоскости (V, U) является множество замкнутых непересекающихся линий, концентрирующихся вокруг точки Е( 1, 1), а решения исходной модели U = Щ) и V = V{t) являются функциями, периодическими во времени. При этом максимум функции U = U(t) не попадает на максимум функции V = V(t ) и наоборот, т. е. колебания численности популяций около своих равновесных решений происходят в разных фазах.
На рис. 6.11 приведены четыре траектории системы дифференциальных уравнений (6.19) на фазовой плоскости OUV, отличающиеся начальными условиями. Одна из траекторий равновесная - это точка Е( 1, 1), которой соответствует решение U(t) = 1, V{t) = 1. Точки (U(t), V(t)) на других трех фазовых траекториях по мере увеличения времени смещаются по часовой стрелке.
Чтобы пояснить действие механизма изменения численности двух популяций, рассмотрим траекторию ABCDA на рис. 6.11. Как видим, на участке АВ и хищников, и жертв мало. Поэтому здесь популяция хищников сокращается из-за недостатка корма, а популяция жертв растет. На участке ВС численность жертв достигает высоких значений, что приводит к росту численности хищников. На участке СА хищников много, и это влечет сокращение численности жертв. При этом после прохождения точки D численность жертв уменьшается на столько, что численность популяции начинает уменьшаться. Цикл замыкается.
Модель «хищник - жертва» - пример структурно неустойчивой модели. Здесь малое изменение правой части одного из уравнений может привести к принципиальному изменению ее фазового портрета.

Рис. 6.11.

Рис. 6.12.
Действительно, если в уравнении динамики жертв учесть внутривидовую конкуренцию, то мы получим систему дифференциальных уравнений:
Здесь при т = 0 популяция жертв развивается согласно логическому закону.
При т Ф 0 ненулевое равновесное решение системы (6.21) при некоторых положительных значениях параметра внутривидовой конкуренции И является устойчивым фокусом, и соответствующие траектории «наматываются» на точку равновесия (рис. 6.12). Если же h = 0, то в этом случае особая точка Е( 1, 1) системы (6.19) является центром, и траектории представляют собой замкнутые линии (см. рис. 6.11).
Замечание. Обычно под моделью «хищник - жертва» понимают модель (6.19), фазовые траектории которой замкнуты. Однако модель (6.21) также является моделью «хищник - жертва», поскольку описывает взаимовлияние хищников и жертв.
Одним из первых приложений модели типа «хищник - жертва» в экономике для исследования циклично изменяющихся процессов является макроэкономическая модель Гудвина, в которой используется непрерывный подход к анализу взаимовлияния уровня занятости и ставки заработной платы.
В работе В.-Б. Занга изложен вариант модели Гудвина, в котором производительность труда и предложение труда растут с постоянными темпами прироста, а коэффициент выбытия фондов равен нулю. Эта модель формально приводит к уравнениям модели «хищник - жертва».
Ниже рассматривается модификация этой модели для случая отличного от нуля коэффициента выбытия фондов.
В модели используются следующие обозначения: L - численность трудящихся; w - средняя ставка заработной платы трудящихся; К - основные производственные фонды (капитал); Y - национальный доход; / - инвестиции; С - потребление; р - коэффициент выбытия фондов; N - предложение труда на рынке рабочей силы; т = Y/K - фондоотдача; а = Y/L - производительность труда; у = L/N - коэффициент занятости; х = C/Y - норма потребления в национальном доходе; К - приращение капитала в зависимости от инвестиций.
Выпишем уравнения модели Гудвина:

где а 0 , b, g, п, N 0 , г - положительные числа (параметры).
Уравнения (6.22) - (6.24) выражают следующее. Уравнение (6.22) - обычное уравнение динамики фондов. Уравнение (6.23) отражает рост ставки заработной платы при высоком уровне занятости (ставка заработной платы растет, если предложение труда невелико) и уменьшение ставки заработной платы при высоком уровне безработицы.
Тем самым уравнение (6.23) выражает закон Филлипса в линейной форме . Уравнения (6.24) означают экспоненциальный рост производительности труда и предложения труда. Предположим также, что С = wL, т. е. вся заработная плата тратится на потребление. Теперь можно преобразовать уравнения модели с учетом равенств:

Преобразуем уравнения (6.22)-(6.27). Имеем:
 где
где
где
![]()
Поэтому динамика переменных в модели Гудвина описывается системой дифференциальных уравнений:

которая формально совпадает с уравнениями классической модели «хищник - жертва». Это значит, что в модели Гудвина тоже возникают колебания фазовых переменных. Механизм колебательной динамики заключается здесь в следующем: при малой заработной плате w потребление низкое, инвестиции большие, и это приводит к росту производства и занятости у. Большая занятость у вызывает рост средней заработной платы w, что приводит к росту потребления и снижению инвестиций, падению производства и снижению занятости у.
Ниже гипотеза о зависимости ставки процента от уровня занятости рассмотренной модели используется при моделировании динамики однопродуктовой фирмы. Оказывается, что в этом случае при некоторых дополнительных допущениях модель фирмы обладает свойством цикличности рассмотренной выше модели «хищник - жертва».
- См.: Вольтерра В. Указ, соч.; Ризнииенко Г. Ю., Рубин А. Б. Указ. соч.
- См.: Занг В.-Б. Синергетическая экономика. М., 2000.
- См.: Пу Т. Нелинейная экономическая динамика. Ижевск, 2000; Тихонов А. Н.Математическая модель // Математическая энциклопедия. Т. 3. М., 1982. С. 574, 575.
Часто представители одного вида (популяции) питаются представителями другого вида.
Модель Лотки – Вольтерры – модель взаимного существования двух популяций типа «хищник – жертва».
Впервые модель «хищник – жертва» была получена А. Лоткой в 1925 году, который использовал ее для описания динамики взаимодействующих биологических популяций. В 1926 году независимо от Лотки аналогичные (к тому же более сложные) модели были разработаны итальянским математиком В. Вольтерра, глубокие исследования которого в области экологических проблем заложили фундамент математической теории биологических сообществ или т. н. математической экологии.
В математической форме предложенная система уравнений имеет вид:
где x – количество жертв, y – количество хищников, t – время, α, β, γ, δ – коэффициенты, которые отражают взаимодействия между популяциями.
Постановка задачи
Рассмотрим закрытое пространство, в котором существуют две популяции – травоядные («жертвы») и хищники. Считается, что животных не ввозят и не вывозят и что еды для травоядных животных достаточно. Тогда уравнение изменения числа жертв (только жертв) примет вид:
где $α$ – коэффициент рождаемости жертв,
$x$– размер популяции жертв,
$\frac{dx}{dt}$ – скорость прироста популяции жертв.
Когда хищники не охотятся, они могут вымирать, значит, уравнение для количества хищников (только хищников) примет вид:
Где $γ$ – коэффициент убыли хищников,
$y$ – размер популяции хищников,
$\frac{dy}{dt}$ – скорость прироста популяции хищников.
При встрече хищников и жертв (частота встреч прямо пропорциональна произведению) хищники уничтожают жертв с коэффициентом, сытые хищники могут воспроизводить потомство с коэффициентом. Таким образом, система уравнений модели примет вид:
Решение задачи
Построим математическую модель совместного существования двух биологических популяций типа «хищник – жертва».
Пусть две биологические популяции совместно обитают в изолированной среде. Среда является стационарной и обеспечивает в неограниченном количестве всем необходимым для жизни один из видов – жертвы. Другой вид – хищник – также обитает в стационарных условиях, но питается только жертвами. В роли хищников могут выступать коты, волки, щуки, лисы, а в роли жертв – куры, зайцы, караси, мыши соответственно.
Для определенности рассмотрим в роли хищников – котов, а в роли жертв – кур.
Итак, куры и коты живут в некотором изолированном пространстве – хозяйственном дворе. Среда предоставляет курам питание в неограниченном количестве, а коты питаются только курами. Обозначим через
$х$ – количество кур,
$у$ – количество котов.
Со временем количество кур и котов меняется, но будем считать $х$ и $у$ непрерывными функциями от времени t. Назовем пару чисел $х, у)$ состоянием модели.
Найдем каким образом изменяется состояние модели $(х, у).$
Рассмотрим $\frac{dx}{dt}$ – скорость изменения количества кур.
Если котов нет, то количество кур возрастает и тем быстрее, чем больше кур. Будем считать зависимость линейной:
$\frac{dx}{dt} a_1 x$,
$a_1$ – коэффициент, который зависит только от условий жизни кур, их естественной смертности и рождаемости.
$\frac{dy}{dt}$ – скорость изменения количества котов (если нет кур), зависит от количества котов y.
Если кур нет, то количество котов уменьшается (у них нет пищи) и они вымирают. Будем считать зависимость линейной:
$\frac{dy}{dt} - a_2 y$.
В экосистеме скорость изменения количества каждого вида также будем считать пропорциональным его количеству, но только с коэффициентом, зависящим от количества особей другого вида. Так, для кур этот коэффициент уменьшается с увеличением количества котов, а для котов возрастает с увеличением количества кур. Будем считать зависимость также линейной. Тогда получим систему дифференциальных уравнений:
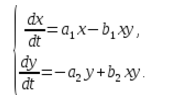
Данная система уравнений называется моделью Вольтерра-Лотки.
a1, a2, b1, b2 – числовые коэффициенты, которые называют параметрами модели.
Как видно, характер изменения состояния модели (x, y) определяется значениями параметров. Изменяя данные параметры и решая систему уравнений модели, можно исследовать закономерности изменения состояния экологической системы.
С помощью программы MATLAB система уравнений Лотки-Вольтерра решается следующим образом:
На рис. 1 представлено решение системы. В зависимости от начальных условий решения разные, чему отвечают разные цвета траекторий.
На рис. 2 представлены те же решения, но с учетом оси времени t (т.е. наблюдается зависимость от времени).
В модели Колмогорова сделано одно существенное допущение: поскольку предполагается, что при то это означает существование в популяции жертв механизмов, регулирующих их численность даже в отсутствие хищников.
К сожалению, подобная формулировка модели не позволяет ответить на вопрос, вокруг которого в последнее время ведется много споров и о котором мы уже упоминали в начале главы: каким образом популяция хищников может оказывать регулирующее воздействие на популяцию жертв с тем, чтобы вся система была устойчива? Поэтому мы вернемся к модели (2.1), в которой механизмы саморегуляции (например, регулирование при помощи внутривидовой конкуренции) в популяции жертв (впрочем, как и в популяции хищников) отсутствуют; поэтому единственным механизмом регуляции численностей, входящих в сообщество видов, являются трофические взаимоотношения между хищниками и жертвами.
Здесь (так что, в отличие от предыдущей модели, Естественно, что решения (2.1) зависят от конкретного вида трофической функции которая, в свою очередь, определяется характером хищничества, т. е. трофической стратегией хищника и защитной стратегией жертвы. Общими для всех этих функций (см. рис. И) являются следующие свойства:
Система (2.1) имеет одну нетривиальную стационарную точку, координаты которой определяются из уравнений
![]()
при естественном ограничении .
Существует еще одна стационарная точка (0, 0), соответствующая тривиальному равновесию. Нетрудно показать, что эта точка - седло, а сепаратрисами являются координатные оси.
Характеристическое уравнение для точки имеет вид
![]()
Очевидно, что для классической вольтерровской модели .
Поэтому величину f можно рассматривать как меру отклонения рассматриваемой модели от вольтерровской.
![]()
стационарная точка - фокус, и в системе появляются колебания; при выполнении противоположного неравенства - узел, и колебания в системе отсутствуют. Устойчивость этого состояния равновесия определяется условием
т. е. существенным образом зависит от вида трофической функции хищника.
Условие (5.5) можно интерпретировать следующим образом: для устойчивости нетривиального равновесия системы хищник - жертва (и тем самым, для существования этой системы) достаточно, чтобы в окрестности этого состояния относительная доля потребляемых хищником жертв возрастала с возрастанием численности жертв. Действительно, доля жертв (от общей их численности), потребляемых хищником, описывается дифференцируемой функцией условие возрастания которой (положительность производной) выглядит как
![]()
Последнее условие, взятое в точке есть не что иное, как условие (5.5) устойчивости равновесия. При непрерывности оно должно выполняться и в некоторой окрестности точки Таким образом, если численности жертв в этой окрестности, то
Пусть теперь трофическая функция V имеет вид, изображенный на рис. 11, а (характерный для беспозвоночных). Можно показать, что для всех конечных значений (поскольку выпукла вверх)
т. е. при любых величинах стационарной численности жертв неравенство (5.5) не выполняется.
Это означает, что в системе с таким видом трофической функции не существует устойчивого нетривиального равновесия. Возможны несколько исходов: либо численности как жертвы, так и хищника неограниченно возрастают, либо (при прохождении траектории вблизи одной из координатных осей) в силу случайных причин численность жертвы или численность хищника станет равной нулю. При гибели жертвы через некоторое время погибнет и хищник, если же сначала погибает хищник, то численность жертвы начнет экспоненциально возрастать. Третий вариант - возникновение устойчивого предельного цикла - невозможен, что легко доказывается.
В самом деле, выражение

в положительном квадранте всегда положительно, если только имеет вид, изображенный на рис. 11, а. Тогда, согласно критерию Дюлака в этой области отсутствуют замкнутые траектории и устойчивого предельного цикла существовать не может.
Итак, можно сделать вывод: если трофическая функция имеет вид, изображенный на рис. 11, а, то хищник не может быть регулятором, обеспечивающим стабильность популяции жертвы и тем самым устойчивость всей системы в целом. Система может быть устойчива только в том случае, когда в популяции жертвы существуют собственные внутренние регулирующие механизмы, например, внутривидовая конкуренция или эпизоотии. Этот вариант регулирования был уже рассмотрен в §§ 3, 4.
Ранее отмечалось, что такой тип трофической функции характерен для хищников-насекомых, «жертвами» которых также обычно являются насекомые. С другой стороны, наблюдения за динамикой многих природных сообществ типа «хищник - жертва», включающих в себя виды насекомых, показывают, что для них характерны колебания очень большой амплитуды и весьма специфического вида.
Обычно после более или менее постепенного роста численности (который может происходить либо монотонно, либо в виде колебаний с возрастающей амплитудой) происходит ее резкое падение (рис. 14), и затем картина повторяется. По-видимому, такой характер динамики численностей видов насекомых может быть объяснен неустойчивостью этой системы при малых и средних значениях численности и действием мощных внутрипопуляционных регуляторов численности при больших ее значениях.

Рис. 14. Динамика численности популяции австралийской листоблошки Cardiaspina albitextura, кормящейся на эвкалиптах. (Из статьи: Clark L. R. The population dynamics of Cardiaspina albitextura.-Austr. J. Zool., 1964, 12, № 3, p. 362-380.)
Если же в систему «хищник - жертва» входят виды, способные к достаточно сложному поведению (например, хищники способны к обучению или жертвы способны находить убежище), то в такой системе возможно существование устойчивого нетривиального равновесия. Доказывается это утверждение весьма просто.
В самом деле, трофическая функция тогда должна иметь вид, изображенной на рис. 11, в. Точка на этом графике - точка касания прямой, проведенной из начала координат, графика трофической функции Очевидно, что в этой точке функция имеет максимум. Также легко показывается, что для всех выполняется условие (5.5). Следовательно, нетривиальное равновесие, в котором численность жертв меньше будет асимптотически устойчиво
Однако мы ничего не можем сказать о том, сколь велика область устойчивости этого равновесия. Например, если существует неустойчивый предельный цикл, то эта область должна лежать внутри цикла. Или другой вариант: нетривиальное равновесие (5.2) неустойчиво, но существует устойчивый предельный цикл; в этом случае также можно говорить о стабильности системы хищник - жертва. Поскольку выражение (5.7) при выборе трофической функции типа рис. 11, в может менять знак при изменении при , то критерий Дюлака здесь не работает и вопрос о существовании предельных циклов остается открытым.

Математическое моделирование биологических процессов началось с создания первых простейших моделей экологической системы.
Допустим, в некотором замкнутом районе живут рыси и зайцы. Рыси питаются только зайцами, а зайцы – растительной пищей, имеющейся в неограниченном количестве. Необходимо найти макроскопические характеристики, описывающие популяции. Такими характеристиками являются число особей в популяциях.
Простейшая модель взаимоотношений популяций хищника и жертвы, основанная на логистическом уравнении роста, названа (как и модель межвидовой конкуренции) по имени ее создателей - Лотки и Вольтерра. Эта модель крайне упрощает исследуемую ситуацию, но все же полезна в качестве отправной точки в анализе системы хищник-жертва.
Предположим, что (1) популяция жертвы существует в идеальной (независимой от плотности) среде, где ее рост может ограничивать только наличие хищника, (2) столь же идеальна среда, в которой существует хищник, рост популяции которого ограничивает лишь обилие жертв, (3) обе популяции размножаются непрерывно согласно экспоненциальному уравнению роста, (4) скорость поедания жертв хищниками пропорциональна частоте встреч между ними, которая, в свою очередь, является функцией плотности популяций. Эти допущения и лежат в основе модели Лотки - Вольтерра.
Пусть в отсутствие хищников популяция жертвы растет экспоненциально:
dN/dt =r 1 N 1
где N -численность, а r, - удельная мгновенная скорость роста популяции жертвы. Если же хищники присутствуют, то они уничтожают особей жертвы со скоростью, которая определяется, во-первых, частотой встреч хищников и жертв, возрастающей по мере увеличения их численностей, и, во-вторых, эффективностью, с которой хищник обнаруживает и ловит свою жертву при встрече. Число жертв, встреченных и съеденных одним хищником N с, пропорционально эффективности охоты, которую мы выразим через коэффициент С 1; численности (плотности) жертвы N и времени, затраченному на поиски Т:
N C =C 1 NT (1)
Из этого выражения легко определить удельную скорость потребления жертв хищником (т.е. число жертв, поедаемых одной особью хищника в единицу времени), которую часто называют также функциональным ответом хищника на плотность популяции жертвы:
В рассматриваемой модели С 1 является константой. Это означает, что число жертв, изъятых хищниками из популяции, линейно возрастает с увеличением ее плотности (так называемый функциональный ответ типа 1). Ясно, что общая скорость поедания жертв всеми особями хищника составит:
![]() (3)
(3)
где Р - численность популяции хищника. Теперь мы можем записать уравнение роста популяции жертвы следующим образом:
При отсутствии жертвы особи хищника голодают и гибнут. Предположим также, что в этом случае численность популяции хищника будет уменьшаться экспоненциально согласно уравнению:
![]() (5)
(5)
где r 2 - удельная мгновенная смертность в популяции хищника.
Если жертвы присутствуют, то те особи хищника, которые смогут их найти и съесть, будут размножаться. Рождаемость в популяции хищника в данной модели зависит только от двух обстоятельств: скорости потребления жертв хищником и эффективности, с которой поглощенная пища перерабатывается хищником в его потомство. Если мы выразим эту эффективность через коэффициент s, то рождаемость составит:
![]()
Поскольку С 1 и s - константы, их произведение - это также константа, которую мы обозначим как С 2 . Тогда скорость роста популяции хищника будет определяться балансом рождаемости и смертности в соответствии с уравнением:
![]() (6)
(6)
Уравнения 4 и 6 вместе образуют модель Лотки-Вольтерра.
Свойства этой модели мы можем исследовать точно так же, как и в случае конкуренции, т.е. построив фазовую диаграмму, на которой численность жертвы отложена по оси ординат, а хищника - по оси абсцисс, и проведя на ней изоклины-линии, соответствующие постоянной численности популяций. С помощью таких изоклин определяют поведение взаимодействующих популяций хищника и жертвы.
Для популяции жертвы: при откуда
Таким образом, поскольку r, и С 1 , - константы, изоклиной для жертвы будет линия, на которой численность хищника (Р) является постоянной, т.е. параллельная оси абсцисс и пересекающая ось ординат в точке Р =r 1 / С 1 . Выше этой линии численность жертвы будет уменьшаться, а ниже- возрастать.
Для популяции хищника:
при откуда
Поскольку r 2 и С 2 - константы, изоклиной для хищника будет линия, на которой численность жертвы (N) является постоянной, т.е. перпендикулярная оси ординат и пересекающая ось абсцисс в точке N = r 2 /С 2 . Слева от нее численность хищника будет уменьшаться, а справа - возрастать.
Если мы рассмотрим эти две изоклины вместе, то легко заметим, что взаимодействие популяций хищника и жертвы имеет циклический характер, поскольку их численности претерпевают неограниченные сопряженные колебания. Когда велико число жертв, численность хищников растет, что приводит к увеличению пресса хищничества на популяцию жертвы и тем самым к снижению ее численности. Это снижение, в свою очередь, ведет к нехватке пищи у хищников и падению их численности, которое вызывает ослабление пресса хищничества и увеличению численности жертвы, что снова приводит к росту популяции жертвы и т.д.
Для данной модели характерна так называемая "нейтральная стабильность", которая означает, что популяции неограниченно долго совершают один и тот же цикл колебаний до тех пор, пока какое-либо внешнее воздействие не изменит их численность, после чего популяции совершают новый цикл колебаний с иными параметрами. Для того, чтобы циклы стали стабильными, популяции должны после внешнего воздействия стремиться вернуться к первоначальному циклу. Такие циклы, в отличие от нейтрально стабильных колебаний в модели Лотки-Вольтерра, принято называть устойчивыми предельными циклами.
Модель Лотки-Вольтерра, тем не менее, полезна тем, что позволяет продемонстрировать основную тенденцию в отношениях хищник-жертва возникновение циклических сопряженных колебаний численности их популяций.
Допущения:
1. Среда однородная.
2. Численность данного вида описывается одной переменной, т.е. мы пренебрегаем возрастными, половыми и генетическими различиями.
3. Пренебрегаем случайными флуктуациями.
4. Взаимодействие мгновенное.
В биологической литературе существует огромное число работ, в которых подобные системы либо наблюдались в природе, либо моделировались на «модельных» популяциях в лабораторных условиях.
Однако их результаты зачастую противоречат друг другу:
− в одних экспериментах наблюдались, на первый взгляд, непонятные явления периодических изменений численности популяций в однородной среде;
− в других наблюдениях системы достаточно быстро разрушались: либо гибнет хищник, а жертва останется, либо гибнет жертва, а вслед за ней хищник.
Построенная в 20-х годах ХХ века Вито Вольтера модель сообщества «хищник-жертва» объясняет многие из этих особенностей.
Это первый успех математической экологии.
При рассмотрении этой системы рассмотрим вопросы устойчивости: условия устойчивости и механизмы устойчивости.
Классическая модель Вольтерра
Численность хищников.
Дополнительные допущения.
1. Единственным лимитирующим фактором, ограничивающим размножение жертв, является давление на них со стороны хищников. Ограниченность ресурсов среды для жертвы не учитывается (как в модели Мальтуса).
2. Размножение хищников ограничивается количеством добытой им пищи (количеством жертв).
− коэффициент естественного прироста жертвы;
− коэффициент естественной смертности хищника;
− количество (биомасса) жертв, потребляемых одним хищником за единицу времени (трофическая функция);
− часть полученной с биомассы энергии, которая расходуется хищником на воспроизводство. Остальная энергии тратится на поддержание основного обмена и охотничьей активности.
Уравнения системы «хищник-жертва»
Функция определяется в экспериментальных работах. К настоящему времени установлено, что эти функции принадлежат к одному из следующих трех типов.
Этот тип характерен для беспозвоночных и некоторых видов хищных рыб.
Трофическая функция с резко выраженным порогом насыщения характерна для хищников - фильтраторов (моллюсков).
Такой тип характерен для позвоночных – организмов, способных к обучению.
При малых значениях численности жертвы почти все жертвы становятся добычей хищника, который всегда голоден и насыщения не наступает. Трофическую функцию можно считать линейной:
Классическая модель Вольтерра:
Начальные условия
Система (2) является автономной, т.к. не имеет в правой части. Изменение состояния системы изображается на фазовой плоскости и является решением уравнения
Найдем точки покоя системы (2).
Нетривиальная точка покоя системы (4) имеет вид
Определим характер точки покоя (5).
Сделаем замену
Раскроем скобки и получим систему
Отбросив нелинейные члены, получим систему
Характеристическое уравнение имеет вид
Корни - чисто мнимые числа. Точка покоя – центр. В исходных переменных фазовые траектории имеют вид
Стрелки указывают направление изменения состояния системы со временем.
Согласно этому движению по траектории численность популяций хищника и жертвы совершают незатухающие периодические колебания, причем колебания численности хищника отстает по фазе от колебаний численности жертвы (на четверть периода).
Фазовый портрет решения имеет вид спирали:
В системе «хищник-жертва» возникают затухающие колебания. Численности жертв и хищников стремятся к своим равновесным значениям (8).
Графики зависимости численностей видов.


