Хищники могут поедать травоядных животных, и также слабых хищников. Хищники обладают широким спектром питания, легко переключаются с одной добычи на другую, более доступную. Хищники часто нападают на слабые жертвы. Поддерживается экологическое равновесие между популяциями жертва-хищник.[ ...]
Если равновесие неустойчиво (предельных циклов нет) или внешний цикл неустойчив, то численности обоих видов, испытывая сильные колебания, уходят из окрестности равновесия. Причем быстрое вырождение (в первой ситуации) наступает при низкой адаптации хищника, т.е. при его высокой смертности (по сравнению со скоростью размножения жертвы). Это означает, что слабый во всех отношениях хищник не способствует стабилизации системы и сам вымирает.[ ...]
Пресс хищников особенно силен, когда в коэволюции хищник - жертва равновесие смещается в сторону хищника и ареал жертвы сужается. Конкурентная борьба тесно связана с нехваткой пищевых ресурсов, она может быть и прямой борьбой, например, хищников за пространство как ресурс, но чаще всего это просто вытеснение вида, которому на данной территории пищи не хватает, видом, которому этого же количества пищи вполне достаточно. Это уже межвидовая конкуренция.[ ...]
 |
И окончательно в системе ’’хищник - жертва”, описываемой моделью (2.7), возникновение диффузионной неустойчивости (при локальной устойчивости равновесия) возможно лишь в том случае, когда естественная смертность хищника возрастает с ростом его численности быстрее, чем линейная функция, и трофическая функция отличается от вольтерровской либо, когда популяция жертвы - это популяция типа Олли.[ ...]
Теоретически в моделях «один хищник - две жертвы» эквивалентное выедание (отсутствие предпочтения того или иного вида жертвы) может повлиять на конкурентное сосуществование видов-жертв лишь в тех местах, где уже существует потенциально устойчивое равновесие . Разнообразие может возрасти только в таких условиях, когда у видов с меньшей конкурентоспособностью скорость роста популяции выше, чем у доминантов. Это позволяет понять ситуацию, когда равномерное выедание ведет к увеличению видового разнообразия растений там, где большее число видов, прошедших отбор на быстрое размножение, сосуществует с видами, эволюция которых направлена на повышение конкурентоспособности .[ ...]
Точно так же выбор жертвы, зависящий от ее плотности, может привести к устойчивому равновесию в теоретических моделях двух конкурирующих видов жертвы, где прежде никакого равновесия не существовало . Для этого хищник должен был бы обладать способностью к функциональным и численным реакциям на изменения плотности жертвы; возможно, однако, что переключение (непропорционально частые нападения на наиболее обильную жертву) будет иметь при этом более важное значение. В самом деле, установлено, что переключение оказывает стабилизирующее влияние в системах «один хищник - п жертв» и представляет собой единственный механизм, способный стабилизировать взаимодействия в тех случаях, когда ниши жертв полностью перекрываются . Такую роль могут играть неспециализированные хищники. Предпочтение более специализированными хищниками доминантного конкурента действует таким же образом, как переключение хищника, и может стабилизировать теоретические взаимодействия в моделях, в которых прежде не существовало равновесия между видами жертвы, при условии что их ниши в какой-то степени разделены .[ ...]
Также- не стабилизирует сообщество и хищник ’’сильный во всех отношениях”, т.е. хорошо адаптированный к данной жертве и с низкой относительной смертностью. При этом система обладает неустойчивым предельным циклом и, несмотря на устойчивость положения равновесия, вырождается в случайной среде (хищник выедает жертву и вследствие этого гибнет). Такая ситуация соответствует медленному вырождению.[ ...]
Таким образом, при хорошей адаптации хищника в окрестности устойчивого равновесия могут возникать неустойчивый и устойчивый циклы, т.е. в зависимости от начальных условий система ’’хищник-жертва” либо стремится к равновесию, либо, колеблясь, уходит от него, либо в окрестности равновесия устанавливаются устойчивые колебания численностей обоих видов.[ ...]
Организмы, которых относят к хищникам, питаются другими организмами, уничтожая свою жертву. Таким образом, среди живых организмов следует выделять еще одну классификационную систему, а именно «хищники» и «жертвы». Отношения между такими организмами складывались на протяжении всей эволюции жизни на нашей планете. Организмы-хищники выступают в роли природных регуляторов численности организмов-жертв. Увеличение численности «хищников» приводит к уменьшению численности «жертв», это, в свою очередь, снижает запасы пищи («жертв») для «хищников», что в целом диктует снижение численности «жертв» и т. д. Таким образом, в биоценозе постоянно происходят колебания численности хищников и жертв, в целом же устанавливается определенное равновесие на какой-то период времени в пределах достаточно устойчивых условий среды.[ ...]
Это в конечном итоге приходит к экологическому равновесию между популяциями хищника и жертвы.[ ...]
Для трофической функции третьего типа состояние равновесия будет устойчивым, если где N - точка перегиба функции (см. рис. 2, в). Это следует из того, что на интервале трофическая функция вогнута и, следовательно, относительная доля потребления хищником жертвы возрастает.[ ...]
Пусть Гг = -Г, т.е. имеет место сообщества типа ’’хищник - жертва”. В этом случае первое слагаемое в выражении (7.4) равно нулю и для выполнения условия устойчивости по вероятности равновесного состояния N требуется, чтобы и второе слагаемое не было положительным.[ ...]
Таким образом, для рассмотренного сообщества типа хищник - жертва можно сделать вывод об асимптотической устойчивости в целом положительного положения равновесия, т. е. для любых начальных данных 1Ч(0)>0 эволюция происходит таким образом, что N(7) - ■ К при условии, что N >0.[ ...]
Так, в однородной среде, не имеющей укрытий для размножения, хищник рано или поздно уничтожает популяцию жертвы и после этого вымирает сам. Волны жизни” (изменений численности хищника и жертвы) следуют друг за другом с постоянным сдвигом по фазе, и в среднем численность как хищника, так и жертвы остается примерно на одном уровне. Длительность периода зависит от скоростей роста обоих видов и от исходных параметров. Для популяции жертвы влияние хищника положительно, так как ее чрезмерное размножение привело бы к краху ее численности. В свою очередь, все механизмы, препятствующие полному истреблению жертвы, способствуют сохранению пищевой базы хищника.[ ...]
Другие модификации могут быть следствием особенностей поведения хищника. Число особей жертвы, которое хищник в состоянии потребить в данное время, имеет свой предел. Эффект насыщения хищника при приближении к этому рубежу показан в табл. 2-4, В. Взаимодействия, описываемые уравнениями 5 и 6, могут иметь устойчивые точки равновесия или обнаруживать циклические колебания. Однако такие циклы отличаются от тех, какие отражены в уравнениях Лотки - Вольтерры 1 и 2. Циклы, передаваемые уравнениями 5 и 6, могут иметь постоянную амплитуду и средние плотности, пока среда постоянна; после того как произошло нарушение, они могут вернуться к прежним амплитудам и средним плотностям. Такие циклы, которые восстанавливаются после нарушений, называются устойчивыми предельными циклами. Взаимодействие зайца и рыси можно считать устойчивым предельным циклом, но это не цикл Лотки - Вольтерры.[ ...]
Рассмотрим возникновение диффузионной неустойчивости в системе ’’хищник -жертва”, но сначапа выпишем условия, обеспечивающие возникновение диффузионной неустойчивости в системе (1.1) при п = 2. Ясно, что равновесие (N , Щ) локально (т.е.[ ...]
Перейдем к интерпретации случаев, связанных с длительным сосуществованием хищника и жертвы. Понятно, что в отсутствие предельных циклов устойчивому равновесию будут соответствовать в случайной среде флуктуации численности, причем их амплитуда будет пропорциональна дисперсии возмущений. Такое явление будет происходить, если хищник имеет высокую относительную смертность и в то же время высокую степень приспособленности к данной жертве.[ ...]
Рассмотрим теперь, как меняется динамика системы с ростом приспособленности хищника, т.е. с убыванием Ъ от 1 до 0. Если приспособленность достаточно низкая, то предельные циклы отсутствуют, а равновесие является неустойчивым. С ростом приспособленности в окрестности этого равновесия возможно появление устойчивого цикла и далее внешнего неустойчивого. В зависимости от начальных условий (соотношения биомассы хищника и жертвы) система может либо терять устойчивость, т.е. уходить из окрестности равновесия, либо в ней будут со временем устанавливаться устойчивые колебания. Дальнейший рост приспособленности делает невозможным колебательный характер поведения системы. Однако при Ъ [ ...]
Пример отрицательной (стабилизирующей) обратной связи - взаимоотношение между хищником и жертвой или функционирование карбонатной системы океана (раствор СОг в воде: СО2 + Н2О -> Н2СОз). Обычно количество углекислоты, растворенное в воде океана, находится в парциальном равновесии с концентрацией углекислого газа в атмосфере. Локальные увеличения углекислоты в атмосфере после извержения вулканов приводят к интенсификации фотосинтеза и поглощению ее карбонатной системой океана. При снижении уровня углекислого газа в атмосфере карбонатная система океана высвобождает СОг в атмосферу. Поэтому концентрация углекислого газа в атмосфере достаточно стабильна.[ ...]
[ ...]
Как отмечает Р.Риклефс (1979), существуют факторы, способствующие стабилизации взаимоотношений в системе “хищник-жертва”: неэффективность хищника, наличие у хищника альтернативных пищевых ресурсов, уменьшение запаздывания в реакции хищника, а также экологические ограничения, налагаемые внешней средой на ту или иную популяцию. Взаимодействия между популяциями хищника и жертвы весьма разнообразны и сложны. Так, если хищники достаточно эффективны, они могут регулировать плотность популяции жертвы, удерживая ее на уровне ниже емкости среды. Через влияние, оказываемое ими на популяции жертвы, хищники воздействуют на эволюцию различных признаков жертвы, что приводит в конечном итоге к экологическому равновесию между популяциями хищника и жертвы.[ ...]
Если выполняется одно из условий: 0 1/2. Если же 6 > 1 (кА [ ...]
Устойчивость биоты и окружающей среды зависит только от взаимодействия растений - автотрофов и растительноядных гетеротрофных организмов. Хищники любых размеров не способны нарушить экологическое равновесие сообщества, так как в естественных условиях они не могут увеличить свою численность при постоянной численности жертв. Хищники не только должны быть сами передвигающимися, но и могут питаться только передвигающимися животными.[ ...]
Никакие другие рыбы не распространены так широко, как щуки. В немногих местах товли в стоячих или проточных водоемах нет давления со стороны щук для сохранения равновесия между жертвой и хищником Только современные искусственные водоемы, в которых щуки являются рыбами нежелательными из-за разведения там других рыб, не заселяются ими целенаправленно. В мире щуки исключительно хорошо представлены. Их ловят по всему северном) полушарию от Соединенных Штатов и Канады в Северной Америке, через Европу до северной части Азии.[ ...]
Еще одна возможность устойчивого сосуществования возникает здесь же, в узком диапазоне сравнительно высокой адаптации. При переходе к неустойчивому режиму с очень ’’хорошим” хищником может возникнуть устойчивый внешний предельный цикл, при котором диссипация биомассы уравновешивается ее притоком в систему (высокой продуктивностью жертвы). Тогда возникает любопытная ситуация, когда наиболее вероятными являются два характерных значения амплитуды случайных колебаний. Одни происходят вблизи равновесия, другие - вблизи предельного цикла, причем возможны более или менее частые переходы между этими режимами.[ ...]
Гипотетические популяции, которые ведут себя в соответствии с векторами на рис. 10.11 А, изображены на рис. 10.11,-Б с помощью графика, показывающего динамику соотношения численностей хищника и жертвы и на рис. 10.11,5 в виде графика динамики численности хищника и жертвы во времени. В популяции жертвы, по мере того как она переходит от равновесия прн низкой плотности к равновесию при высокой и возвращается назад, происходит «вспышка» численности. И эта вспышка численности не является следствием столь же выраженного изменения в окружающей среде. Напротив, это изменение численности порождено самим воздействием (при небольшом уровне «шума» в среде) и оно, в частности, отражает существование нескольких равновесных состояний. Сходные рассуждения можно использовать для объяснения более сложных случаев динамики численности в природных популяциях.[ ...]
Важнейшим свойством экосистемы является ее устойчивость, сбалансированность обмена и происходящих в ней процессов. Способность популяций или экосистемы поддерживать устойчивое динамическое равновесие в изменяющихся условиях среды называется гомеостазом (homoios - тот же, подобный; stasis - состояние). В основе гомеостаза лежит принцип обратной связи. Для поддержания равновесия в природе не требуется внешнего управления. Пример гомеостаза - субсистема «хищник-жертва», в которой регулируются плотность популяций хищника и жертвы.[ ...]
Природная экосистема (биогеоценоз) устойчиво функционирует при постоянном взаимодействии ее элементов, круговороте веществ, передаче химической, энергетической, генетической и другой энергии и информации по цепям-каналам. Согласно принципу равновесия любая естественная система с проходящим через нее потоком энергии и информации имеет тенденцию к развитию устойчивого состояния. При этом устойчивость экосистем обеспечивается автоматически за счет механизма обратной связи. Обратная связь заключается в использовании получаемых данных от управляемых компонентов экосистем для внесения корректив управляющими компонентами в процесс. Рассмотренные выше взаимоотношения «хищник» - «жертва» в данном контексте можно описать несколько подробнее; так, в водной экосистеме хищные рыбы (щука в пруду) поедают другие виды рыб-жертвы (карась); если численность карася будет увеличиваться -это пример положительной обратной связи; щука, питаясь карасем, снижает его численность-это пример отрицательной обратной связи; при росте числа хищников снижается число жертв, и хищник, испытывая недостаток пищи, также снижает рост своей популяции; в конце концов в рассматриваемом пруду устанавливается динамическое равновесие в численности и щуки, и карася. Постоянно поддерживается равновесие, которое исключало бы исчезновение любого звена трофической цепи (рис. 64).[ ...]
Переходим к наиболее важному обобщению, а именно что отрицательные взаимодействия со временем становятся менее заметными, если экосистема достаточно стабильна и ее пространственная структура обеспечивает возможность взаимного приспособления популяций. В модельных системах типа хищник- жертва, описываемых уравнением Лотки-Вольтерры, если в уравнение не введены дополнительные члены, характеризующие действие факторов самоограничения численности, то колебания происходят непрерывно и не затухают (см. Левонтин, 1969). Пиментел (1968; см. также Пиментел и Стоун, 1968) экспериментально показал, что такие дополнительные члены могут отражать взаимные адаптации или генетическую обратную связь. Когда же новые культуры создавали из особей, ранее на протяжении двух лет совместно существовавших в культуре, где их численность подвергалась значительным колебаниям, оказалось, что у них выработался экологический гомеостаз, при котором каждая из популяций была «подавлена» другой в такой степени, что оказалось возможным их сосуществование при более стабильном равновесии.
Федеральное агентство по образованию
Государственное образовательное учреждение
высшего профессионального образования
«Ижевский государственный технический университет»
Факультет «Прикладная математика»
Кафедра «Математическое моделирование процессов и технологий»
по дисциплине «Дифференциальные уравнения»
Тема: «Качественное исследование модели хищник-жертва»
Ижевск 2010
ВВЕДЕНИЕ
1. ПАРАМЕТРЫ И ОСНОВНОЕ УРАВНЕНИЕ МОДЕЛИ «ХИЩНИК-ЖЕРТВА»
2.2 Обобщенные модели Вольтера типа «хищник-жертва».
3. ПРАКТИЧЕСКОЕ ПРИМЕНЕНИЕ МОДЕЛИ «ХИЩНИК-ЖЕРТВА»
ЗАКЛЮЧЕНИЕ
СПИСОК ЛИТЕРАТУРЫ
ВВЕДЕНИЕ
В настоящее время задачи экологии имеют первостепенное значение. Важным этапом решения этих задач является разработка математических моделей экологических систем.
Одной из основных задач экологии па современном этапе является изучение структуры и функционирования природных систем, поиск общих закономерностей. Большое влияние на экологию оказала математика, способствующая становлению математической экологии, особенно такие её разделы, как теория дифференциальных уравнений, теория устойчивости и теория оптимального управления.
Одной из первых работ в области математической экологии была работа А.Д. Лотки (1880 - 1949), который первый описал взаимодействие различных популяций, связанных отношениями хищник - жертва. Большой вклад в исследование модели хищник -жертва внесли В. Вольтерра (1860 - 1940), В.А. Костицин (1883-1963) В настоящее время уравнения описывающие взаимодействие популяций, называются уравнениями Лотки - Вольтерра.
Уравнения Лотки - Вольтерра описывают динамику средних величин - численности популяции. В настоящее время на их основе построены более общие модели взаимодействия популяций, описываемые интегро-дифференциальными уравнениями, исследуются управляемые модели хищник - жертва.
Одной из важных проблем математической экологии является проблема устойчивости экосистем, управления этими системами. Управление может осуществляться с целью перевода системы из одного устойчивого состояния в другое, с целью её использования или восстановления.
1. ПАРАМЕТРЫ И ОСНОВНОЕ УРАВНЕНИЕ МОДЕЛИ ХИЩНИК-ЖЕРТВА
Попытки математического моделирования динамики как отдельных биологических популяций, так и сообществ, включающих взаимодействующие популяции различных видов, предпринимались давно. Одна из первых моделей роста изолированной популяции (2.1) была предложена еще в 1798 г. Томасом Мальтусом:
, (1.1)Данная модель задается следующими параметрами:
N - численность популяции;
- разность между коэффициентами рождаемости и смертности.Интегрируя это уравнение получаем:
, (1.2)где N(0) – численность популяции в момент t = 0. Очевидно, что модель Мальтуса при
> 0 дает бесконечный рост численности, что никогда не наблюдается в природных популяциях, где ресурсы, обеспечивающие этот рост, всегда ограничены. Изменения численности популяций растительного и животного мира нельзя описывать простым законом Мальтуса, на динамику роста влияют многие взаимосвязанные причины – в частности, размножение каждого вида саморегулируется и видоизменяется так, чтобы этот вид сохранялся в процессе эволюции.Математическим описанием этих закономерностей занимается математическая экология – наука об отношениях растительных и животных организмов и образуемых ими сообществ между собой и с окружающей средой.
Наиболее серьезное исследование моделей биологических сообществ, включающих в себя несколько популяций различных видов, было проведено итальянским математиком Вито Вольтерра:
, - численность популяции; - коэффициенты естественного прироста (или смертности) популяции; - коэффициенты межвидового взаимодействия. В зависимости от выбора коэффициентов модель описывает либо борьбу видов за общий ресурс, либо взаимодействие типа хищник - жертва, когда один вид является пищей для другого. Если в работах других авторов основное внимание уделялось построению различных моделей, то В. Вольтерра провел глубокое исследование построенных моделей биологических сообществ. Именно с книги В. Вольтерра, по мнению многих ученых, началась современная математическая экология.2. КАЧЕСТВЕННОЕ ИССЛЕДОВАНИЕ ЭЛЕМЕНТАРНОЙ МОДЕЛИ «ХИЩНИК-ЖЕРТВА»
2.1 Модель трофического взаимодействия по типу «хищник-жертва»
Рассмотрим модель трофического взаимодействия по типу «хищник-жертва», построенную В. Вольтерром. Пусть имеется система, состоящая из двух видов, из которых один поедает другой.
Рассмотрим случай, когда один из видов является хищником, а другой - жертвой, и будем считать, что хищник питается только жертвой. Примем следующую простую гипотезу:
- коэффициент прироста жертвы; - коэффициент прироста хищника; - численность популяции жертвы; - численность популяции хищника; - коэффициент естественного прироста жертвы; - скорость потребления жертвы хищником; - коэффициент смертности хищника в отсутствие жертвы; - коэффициент «переработки» хищником биомассы жертвы в собственную биомассу.Тогда динамика численности популяций в системе хищник - жертва будет описываться системой дифференциальных уравнений (2.1):
(2.1)где все коэффициенты положительные и постоянные.
Модель имеет равновесное решение (2.2):
(2.2)По модели (2.1) доля хищников в общей массе животных выражается формулой (2.3):
(2.3)Анализ устойчивости состояния равновесия по отношению к малым возмущениям показал, что особая точка (2.2) является «нейтрально» устойчивой (типа «центр»), т. е. любые отклонения от равновесия не затухают, но переводят систему в колебательный режим с амплитудой, зависящей от величины возмущения. Траектории системы на фазовой плоскости
имеют вид замкнутых кривых, расположенных на различных расстояниях от точки равновесия (рис. 1).Рис. 1 – Фазовый «портрет» классической вольтерровой системы «хищник-жертва»
Разделив первое уравнение системы (2.1) на второе, получим дифференциальное уравнение (2.4) для кривой на фазовой плоскости
. (2.4)Интегрируя данное уравнение получим:
(2.5) - постоянная интегрирования, гдеНесложно показать, что движение точки по фазовой плоскости будет происходить только в одну сторону. Для этого удобно сделать замену функций
и , перенеся начало координат на плоскости в стационарную точку (2.2) и введя затем полярные координаты: (2.6)В таком случае, подставив значения системы (2.6) в систему (2.1), будем иметь.
Модель ситуации типа «хищник-жертва»
Рассмотрим математическую модель динамики сосуществования двух биологических видов (популяций), взаимодействующих между собой по типу «хищник-жертва» (волки и кролики, щуки и караси и т.д.), называемую моделью Волътера-Лотки. Впервые она была получена А. Лоткой (1925 г.), А чуть позже и независимо от Лотки аналогичные и более сложные модели были разработаны итальянским математиком В. Вольтерра (1926 г.), работы которого фактически заложили основы так называемой математической экологии.
Пусть есть два биологических вида, которые совместно обитают в изолированной среде. При этом предполагается:
- 1. Жертва может найти достаточно пищи для пропитания;
- 2. При каждой встрече жертвы с хищником последний убивает жертву.
Будем для определённости называть их карасями и щуками. Пусть
состояние системы определяется величинами x(t) и y(t) - количеством карасей и щук в момент г. Чтобы получить математические уравнения, которые приближенно описывают динамику (изменение во времени) популяции, поступим следующим образом.
Как и в предыдущей модели роста популяции (см. п. 1.1) для жертв имеем уравнение
где а > 0 (рождаемость превышает смертность)
Коэффициент а прироста жертв зависит от количества хищников (уменьшается с их увеличением). В простейшем случае а- а - fjy (а>0,р>0). Тогда для численности популяции жертв имеем дифференциальное уравнение
Для популяции хищников имеем уравнение

где b >0 (смертность превышает рождаемость).
Коэффициент b убывания хищников уменьшается, если имеются жертвы, которыми можно питаться. В простейшем случае можно принять b - у -Sx {у > 0, S > 0). Тогда для численности популяции хищников получим дифференциальное уравнение

Таким образом, уравнения (1.5) и (1.6) представляют собой математическую модель рассматриваемой задачи взаимодействия популяций. В этой модели переменные х,у - состояние системы, а коэффициенты характеризуют ее структуру. Нелинейная система (1.5), (1.6) и есть модель Вольтера-Лотки.
Уравнения (1.5) и (1.6) следует дополнить начальными условиями - заданными значениями начальных популяций.
Проведем теперь анализ построенной математической модели.
Посгроим фазовый портрет системы (1.5), (1.6) (по смыслу задачи х > 0, v >0). Разделив уравнение (1.5) на уравнение (1.6), получим уравнение с разделяющимися переменными
Игшлрируя это уравнение, будем иметь
Соотношение (1.7) даёт уравнение фазовых траекторий в неявном виде. Система (1.5), (1.6) имеет стационарное состояние определяемое из

Из уравнений (1.8) получим (т.к. л* Ф 0, у* Ф 0)
Равенства (1.9) определяют на фазовой плоскости положение равновесия (точку О) (Рис. 1.6).

Направление движения по фазовой траектории можно определить из таких соображений. Пусть карасей мало. г.е. х ~ 0, тогда из уравнения (1.6) у
Все фазовые траектории (за исключением точки 0) замкнутые кривые, охватывающие положение равновесия. Состоянию равновесия соответствует неизменное количество х« и у« карасей и щук. Караси размножаются, щуки их едят, вымирают, но число тех и дрч их не меняется. "Замкнутым фазовым траекториям соответствует периодическое изменение численности карасей и щук. Причём то, по какой траектории движется фазовая точка, зависит от начальных условий. Рассмотрим, как меняется состояние вдоль фазовой траектории. Пусть точка находится в положении А (рис. 1.6). Здесь карасей мало, щук много; щукам есть нечего, и они постепенно вымирают и почти
совсем исчезают. Но и количество карасей тоже уменьшается почти до нуля и
только потом, когда щук стало меньше, чем у , начинается прирост количества карасей; скорость их прироста увеличивается и их число увеличивается - так происходит примерно до точки В. Но увеличение числа карасей приводит к торможению процесса вымирания шук и их число начинает расти (пищи стало больше) - участок ВС. Далее щук много, они едят карасей и почти всех съедают (участок CD). После этого щуки снова начинают вымирать и процесс повторяется с периодом примерно в 5-7 лет. На рис. 1.7 качественно построены кривые изменения численности карасей и щук в зависимости от времени. Максимумы кривых чередуются, причём максимумы численности щук отстают от максимумов карасей.

Такое поведение характерно для различных систем типа хищник - жертва. Проведем теперь интерпретацию полученных результатов.
Несмотря на то, что рассмотренная модель является простейшей и в действительности всё происходит гораздо сложнее, она позво.чила объяснить кое-что из загадочного, чго есть в природе. Понятны рассказы рыболовов о периодах, когда «щуки сами прыгают в руки», получила объяснение периодичность протекания хронических болезней и т.д.
Отметим еще один интересный вывод, который можно сделать из Рис. 1.6. Если в точке Р происходит быстрый отлов щук (в другой терминологии - отстрел волков), то система «перепрыгивает » в точку Q, и дальнейшее движение происходит по замкнутой траектории меньшего размера, что интуитивно ожидаемо. Если же уменьшить число щук в точке R, то система перейдет в точку S, и дальнейшее движение будет происходить по траектории большего размера. Амплитуды колебаний увеличатся. Это противоречит интуиции, но как раз объясняет такое явление: в результате отстрела волков их численность увеличивается со временем. Таким образом, важным в этом случае является выбор момента отстрела.
Предположим, что две популяции насекомых (например, тля и божья коровка, которая есть тлю) находились в естественном равновесии х-х*,у = у* (точка О на Рис. 1.6). Рассмотрим влияние разового применения инсектицида, который убивает х> 0 из жертв и у > 0 из хищников, не уничтожая их полностью. Уменьшение численности обеих популяций приводит к тому, что изображающая точка из положения О «перескочит» ближе к началу координат, где х > 0, у 0 (Рис. 1.6) Отсюда следует, что в результате действия инсектицида, призванного уничтожать жертв (тлю), число жертв (тли) увеличивается, а число хищников (божьих коровок) уменьшается. Получается, что численность хищников может стать настолько малой, что им будет фозить полное исчезновение но другим причинам (засуха, болезни и т.д.). Таким образом, применение инсектицидов (если только они не уничтожают вредных насекомых практически полностью) в конечном счёте приводит к увеличению популяции тех насекомых, численность которых находилась под контролем других насекомых-хищников. Такие случаи описаны в книгах по биологии.
В общем случае коэффициент прироста количества жертв а зависит и от Л" и от у: а = а(х, у) (из-за наличия хищников и из-за ограничений на пищу).
При малом изменении модели (1.5), (1.6) к правым частям уравнений добавляются малые члены (учитывающие, например, конкуренцию карасей за пищу и щук за карасей)
здесь 0 f.i « 1.
В таком случае вывод о периодичности процесса (возвращении системы к исходному состоянию), справедливый для модели (1.5), (1.6), теряет силу. В зависимости от вида малых поправок/ и g возможны ситуации, показанные на Рис. 1.8.

В случае (1) равновесное состояние О устойчиво. При любых других начальных условиях через достаточно большое время устанавливается именно оно.
В случае (2) система «идёт в разнос». Стационарное состояние неустойчиво. Такая система в конце концов попадает в такую область значений х и у, что модель перестаёт быть применимой.
В случае (3) в системе с неустойчивым стационарным состоянием О устанавливается с течением времени периодический режим. В отличие от исходной модели (1.5), (1.6) в этой модели установившийся периодический режим не зависит от начальных условий. Первоначально малое отклонение от стационарного состояния О приводит не к малым колебаниям около О , как в модели Вольтерра-Лотки, а к колебаним вполне определённой (и не зависящей от малости отклонения) амплитуды.
В.И. Арнольд называет модель Вольтерра-Лотки жесткой, т.к. её малое изменение может привести к выводам, отличным от приведенных выше. Для суждения о том, какая из ситуаций, указанных на Рис. 1.8, реализуется в данной системе, совершенно необходима дополнительная информация о системе (о виде малых поправок/ и g ).
Часто представители одного вида (популяции) питаются представителями другого вида.
Модель Лотки – Вольтерры – модель взаимного существования двух популяций типа «хищник – жертва».
Впервые модель «хищник – жертва» была получена А. Лоткой в 1925 году, который использовал ее для описания динамики взаимодействующих биологических популяций. В 1926 году независимо от Лотки аналогичные (к тому же более сложные) модели были разработаны итальянским математиком В. Вольтерра, глубокие исследования которого в области экологических проблем заложили фундамент математической теории биологических сообществ или т. н. математической экологии.
В математической форме предложенная система уравнений имеет вид:
где x – количество жертв, y – количество хищников, t – время, α, β, γ, δ – коэффициенты, которые отражают взаимодействия между популяциями.
Постановка задачи
Рассмотрим закрытое пространство, в котором существуют две популяции – травоядные («жертвы») и хищники. Считается, что животных не ввозят и не вывозят и что еды для травоядных животных достаточно. Тогда уравнение изменения числа жертв (только жертв) примет вид:
где $α$ – коэффициент рождаемости жертв,
$x$– размер популяции жертв,
$\frac{dx}{dt}$ – скорость прироста популяции жертв.
Когда хищники не охотятся, они могут вымирать, значит, уравнение для количества хищников (только хищников) примет вид:
Где $γ$ – коэффициент убыли хищников,
$y$ – размер популяции хищников,
$\frac{dy}{dt}$ – скорость прироста популяции хищников.
При встрече хищников и жертв (частота встреч прямо пропорциональна произведению) хищники уничтожают жертв с коэффициентом, сытые хищники могут воспроизводить потомство с коэффициентом. Таким образом, система уравнений модели примет вид:
Решение задачи
Построим математическую модель совместного существования двух биологических популяций типа «хищник – жертва».
Пусть две биологические популяции совместно обитают в изолированной среде. Среда является стационарной и обеспечивает в неограниченном количестве всем необходимым для жизни один из видов – жертвы. Другой вид – хищник – также обитает в стационарных условиях, но питается только жертвами. В роли хищников могут выступать коты, волки, щуки, лисы, а в роли жертв – куры, зайцы, караси, мыши соответственно.
Для определенности рассмотрим в роли хищников – котов, а в роли жертв – кур.
Итак, куры и коты живут в некотором изолированном пространстве – хозяйственном дворе. Среда предоставляет курам питание в неограниченном количестве, а коты питаются только курами. Обозначим через
$х$ – количество кур,
$у$ – количество котов.
Со временем количество кур и котов меняется, но будем считать $х$ и $у$ непрерывными функциями от времени t. Назовем пару чисел $х, у)$ состоянием модели.
Найдем каким образом изменяется состояние модели $(х, у).$
Рассмотрим $\frac{dx}{dt}$ – скорость изменения количества кур.
Если котов нет, то количество кур возрастает и тем быстрее, чем больше кур. Будем считать зависимость линейной:
$\frac{dx}{dt} a_1 x$,
$a_1$ – коэффициент, который зависит только от условий жизни кур, их естественной смертности и рождаемости.
$\frac{dy}{dt}$ – скорость изменения количества котов (если нет кур), зависит от количества котов y.
Если кур нет, то количество котов уменьшается (у них нет пищи) и они вымирают. Будем считать зависимость линейной:
$\frac{dy}{dt} - a_2 y$.
В экосистеме скорость изменения количества каждого вида также будем считать пропорциональным его количеству, но только с коэффициентом, зависящим от количества особей другого вида. Так, для кур этот коэффициент уменьшается с увеличением количества котов, а для котов возрастает с увеличением количества кур. Будем считать зависимость также линейной. Тогда получим систему дифференциальных уравнений:
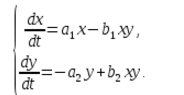
Данная система уравнений называется моделью Вольтерра-Лотки.
a1, a2, b1, b2 – числовые коэффициенты, которые называют параметрами модели.
Как видно, характер изменения состояния модели (x, y) определяется значениями параметров. Изменяя данные параметры и решая систему уравнений модели, можно исследовать закономерности изменения состояния экологической системы.
С помощью программы MATLAB система уравнений Лотки-Вольтерра решается следующим образом:
На рис. 1 представлено решение системы. В зависимости от начальных условий решения разные, чему отвечают разные цвета траекторий.
На рис. 2 представлены те же решения, но с учетом оси времени t (т.е. наблюдается зависимость от времени).
Допущения:
1. Среда однородная.
2. Численность данного вида описывается одной переменной, т.е. мы пренебрегаем возрастными, половыми и генетическими различиями.
3. Пренебрегаем случайными флуктуациями.
4. Взаимодействие мгновенное.
В биологической литературе существует огромное число работ, в которых подобные системы либо наблюдались в природе, либо моделировались на «модельных» популяциях в лабораторных условиях.
Однако их результаты зачастую противоречат друг другу:
− в одних экспериментах наблюдались, на первый взгляд, непонятные явления периодических изменений численности популяций в однородной среде;
− в других наблюдениях системы достаточно быстро разрушались: либо гибнет хищник, а жертва останется, либо гибнет жертва, а вслед за ней хищник.
Построенная в 20-х годах ХХ века Вито Вольтера модель сообщества «хищник-жертва» объясняет многие из этих особенностей.
Это первый успех математической экологии.
При рассмотрении этой системы рассмотрим вопросы устойчивости: условия устойчивости и механизмы устойчивости.
Классическая модель Вольтерра
Численность хищников.
Дополнительные допущения.
1. Единственным лимитирующим фактором, ограничивающим размножение жертв, является давление на них со стороны хищников. Ограниченность ресурсов среды для жертвы не учитывается (как в модели Мальтуса).
2. Размножение хищников ограничивается количеством добытой им пищи (количеством жертв).
− коэффициент естественного прироста жертвы;
− коэффициент естественной смертности хищника;
− количество (биомасса) жертв, потребляемых одним хищником за единицу времени (трофическая функция);
− часть полученной с биомассы энергии, которая расходуется хищником на воспроизводство. Остальная энергии тратится на поддержание основного обмена и охотничьей активности.
Уравнения системы «хищник-жертва»
Функция определяется в экспериментальных работах. К настоящему времени установлено, что эти функции принадлежат к одному из следующих трех типов.
Этот тип характерен для беспозвоночных и некоторых видов хищных рыб.
Трофическая функция с резко выраженным порогом насыщения характерна для хищников - фильтраторов (моллюсков).
Такой тип характерен для позвоночных – организмов, способных к обучению.
При малых значениях численности жертвы почти все жертвы становятся добычей хищника, который всегда голоден и насыщения не наступает. Трофическую функцию можно считать линейной:
Классическая модель Вольтерра:
Начальные условия
Система (2) является автономной, т.к. не имеет в правой части. Изменение состояния системы изображается на фазовой плоскости и является решением уравнения
Найдем точки покоя системы (2).
Нетривиальная точка покоя системы (4) имеет вид
Определим характер точки покоя (5).
Сделаем замену
Раскроем скобки и получим систему
Отбросив нелинейные члены, получим систему
Характеристическое уравнение имеет вид
Корни - чисто мнимые числа. Точка покоя – центр. В исходных переменных фазовые траектории имеют вид
Стрелки указывают направление изменения состояния системы со временем.
Согласно этому движению по траектории численность популяций хищника и жертвы совершают незатухающие периодические колебания, причем колебания численности хищника отстает по фазе от колебаний численности жертвы (на четверть периода).
Фазовый портрет решения имеет вид спирали:
В системе «хищник-жертва» возникают затухающие колебания. Численности жертв и хищников стремятся к своим равновесным значениям (8).
Графики зависимости численностей видов.


